动物的采食对策包括动物对采食时间、食物种类和采食地的选择(Nagy , 1994; Julien2Laferrière , 措施。1999)。MacArthur et al. (1966)提出了动物利用斑块环境的采食对策,Charnov(1976)、Stephenet al. (1986)提出了优化采食理论。他们的研究涉及夜猴 ( Aotus trivirgatus) 、美洲狮 ( Pumaconcolor) 和美洲野牛 ( Bison bison) 的采食时间、食物种类和采食地的选择及其影响因素,促进了采食对策理论的发展(Steuter et al. , 1995; Wright、1989; Kunkel et al. , 1999)。
普氏原羚 ( Procapra przewalskii)现在仅分布于环青海湖的青海省刚察、海晏和共和县。20 世纪 60 年代 , 普氏原羚曾是当地一种常见动物 , 现仅存 300 只左右 (蒋志刚等 , 1995; 2001) 。导致濒危的原因很多 , 如栖息地被破坏、过度捕猎、食物匮乏和人类活动增加等。在栖息地被破坏的势头得到遏制,捕猎得到有效控制后,季节性的食物匮乏和人类活动的增加成为普氏原羚生存面临的主要威胁,对这两方面的适应是普氏原羚面临的首要问题。通过研究普氏原羚的采食时间、食物选择以及采食地选择,探讨了普氏原羚对食物资源季节性变化和人类活动增加的适应,提出了普氏原羚的保护措施。
1研究地点和研究方法
1.1研究地点
研究地点位于青海省海晏县和共和县,东经99°50′~100°46′,北纬36°41′~37°55′,海拔3 036 m~3 226 m。研究地区面积为 1 000 km 左右,人口 2 100 人左右,其中可利用草地和农业用地面积占7516 %。气候条件已有报道 (李迪强等 , 1999) 。该地区有芨芨草 ( Achnatherum splendens) 草原、狼毒 (Stellera chamaejasme)草原、沙蒿(Artemisia desertorum)灌丛、疏花针茅)。沙丘是普氏原羚的重要栖息场所。芨芨草草原是普氏原羚分布区主要的植被类型,优势 植物有芨芨草、赖草 ( Leymussecalinus) 、冰草 ( Agropyron cristatum) 和青藏苔草 ( Carex mooscroftii) 等,这些植物都是优良的牧草,普氏原羚亦经常采食这些植物;狼毒草原分布在沙丘的外围,以狼毒、碱蒿 ( Artemisia anethifolia) 、萎陵菜 ( Potentilla chinensis) 等植物为建群种; 沙蒿灌丛分布在沙丘草原交界处, 建群植物是沙蒿。沙蒿是草原上返青生长较早、生长迅速的植物;疏花针茅草原主要分布在沙丘和草原的交界处,主要植物有针茅 ( Stipa spp.) 、披针叶黄华 ( Thermopsis lanceolata) 、唐古特铁线莲( Clematis tangutica) 等。沙生针茅草原主要分布在沙丘内,优势植物有青藏苔草 ( Carexmooscroftii) 和青海野青茅 ( Deyeuxia kdconocia) ;马蔺草甸主要分布于湖边等生境,建群植物是马蔺( Iris lactea) 。此外,在分布区内还分布有沙地柏(Sabina vulgaris)灌丛和金露梅 ( Potentillaparrifolia) 灌丛,这两种植被类型都不能为普氏原羚提供食物。
湖东种羊场 2000 年有 36 900 只绵羊、8 900只种绵羊、1 763 头牦牛和 222 匹马。研究地区为牧区,草场分包到户,牧民用铁丝将草场围起来,自主经营。牧民实行季节轮牧制度。夏秋牧场位于研究地区的山上,没有普氏原羚的分布。普氏原羚在冬春季牧场活动。8 月份普氏原羚活动区没有大群放牧的家畜。11 月份到次年 5 月份,大群放牧的家畜回归冬春牧场,家畜在普氏原羚活动区采食,与普氏原羚竞争食物,普氏原羚食物匮乏。
1.2研究方法
从 1998 年 8 月到 2001 年 6 月,我们运用粪样显微组织学分析法研究了普氏原羚的食物组成(William,1969; 陈化鹏等,1989)。在野外收集了当地常见的植物,做成标准样本玻片。1998年 8、9、10 和 11 月,1999 年 3、5、6 和 8 月,2000 年5月和 6 月,每月随机收集至少 20 个普氏原羚的粪样。粪样于 60 ℃烘干 30 h 后保存。显微组织学分析时,每个粪样取 3 粒,构成混合粪样,研碎,用番红染色,每个样品做 10 个玻片,每个玻片利用频率转换法在显微镜下放大 100 倍进行观察,区分50 个视野,与标准样本玻片进行对照记录普氏原羚粪样中的植物组成。为了研究植物生物量对普氏原羚采食对策的影响,我们对湖东 - 克图普氏原羚分布区内芨芨草草原 (15 m ×0.15 m)、狼毒草原(0.15 m ×0.15 m)、疏花针茅草原(0.15 m ×0.15 m)、沙生针茅草原(0.15 m ×0.15 m)、沙蒿灌丛(1 m ×1 m)和马蔺草甸(0.15 m ×0.15 m)中 (括号内为取样面积) 选取3 个样方 , 各样方间隔 50 m , 剪取样方内植物的地上部分 , 剪取沙蒿灌丛普氏原羚取食部分。植物样品于 60 ℃烘干至恒重 , 称量并记录每个样方内的植物总重量。
此外 , 作者每天 6: 00~22: 00 沿固定路线(40 km左右)在草原和沙丘交错带以 10 km/ h 左右的速度调查。记录普氏原羚群体大小、群体组成、行为、出现地的植被类型、草原围栏情况、隐蔽情况、人为干扰强度、与公路距离、与沙丘距离、与农业用地距离等。利用目标动物取样法(Focal2animal sampling method) 记录在不受惊扰条件下 , 处于群体最外面普氏原羚个体的行为 , 计算采食行为占所有行为的比例。
利用单因素方差技术分析不同月份间普氏原采食活动频次 , 比较食物资源量和人为干扰对普原羚采食活动的影响。高的人类活动强度是指在氏原羚活动区从 8: 00 到 18: 00 一直有大群放的家畜 , 标准为在视野开阔处可以见到牧民或者群采食的家畜。否则 , 定为人类活动强度比较低利用 Kolmogorov2Smirnov 检验和配对 t 检验分普氏原羚的食物组成的季节性差异。采用选择指Ei研究普氏原羚对采食生境的选择 (Lechowic1982) 。
2结 果
2.1食物资源
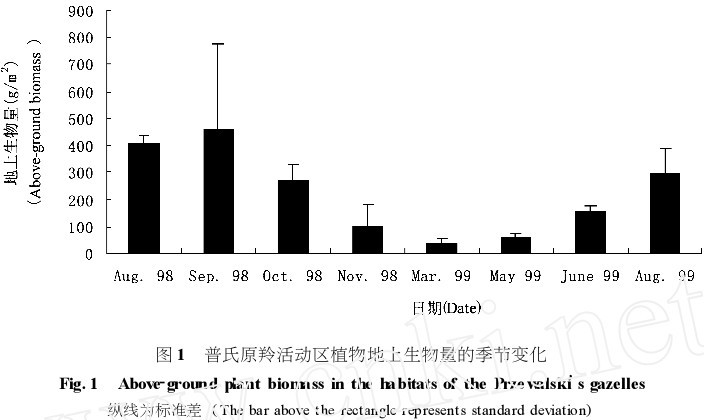
植物地上生物量季节性变化大 (图 1) 。9月份 , 植物地上生物量干重为 45713 g/ m , 并且有家畜的分布。3 月植物生物量最低 , 植物地上植物量干重为 3610 g/ m,仅为 9 月生物量的 719 %。从11月到来年5月,植物地上生物量均低于 9月植物地上生物量的 10 %。
2.2 采食时间
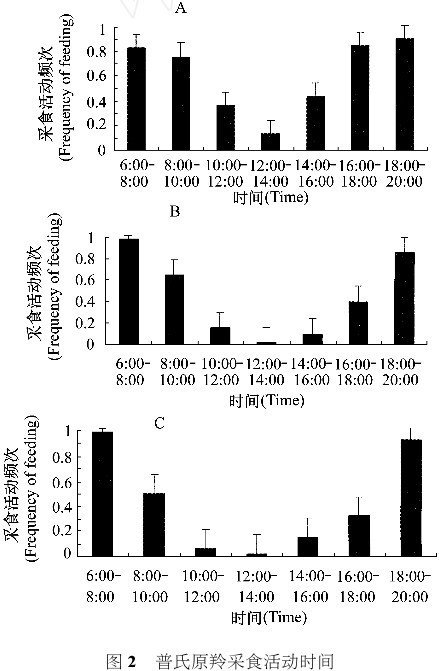
调查中,共发现普氏原羚 942 只次,行为记录持续时间 4 104 min,其中记录采食时间行为2 08418 min。从 6: 00 到 20: 00 普氏原羚采食活动占 4010 %~6516 %(图2) 。除6: 00~8: 00时间段外 , 8 月普氏原羚采食活动频次高于 6 月和10月。8 月人为干扰强度小时 , 普氏原羚一天中采食时间分配较均衡 , 10: 00 到 16: 00 采食活动明显。对普氏原羚采食活动频次进行单因素方差分析表明 , 8 月与 6 月和 10 月采食活动频次差异极显著 ( P < 0101) 。3 月、5 月和 11 月从 10: 00到14: 00 普氏原羚采食活动少。6 月和 10 月相比较、普氏原羚从 10: 00 到 16: 00 的采食活动频次差异极显著( P < 0101) 。食物资源明显影响了普氏原羚采食活动。
2.3 食物组成
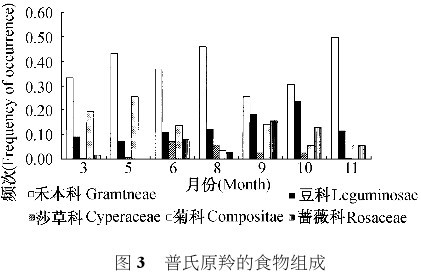
我们采集到植物 22 科 79 种。粪样分析发现:普氏原羚共取食 16 科 50 种植物。禾本科是普氏原羚采食的主要植物类群,占可识别植物碎片的3315 %~6814 %,其次是豆科、菊科、蔷薇科和莎草科。5 科植物在普氏原羚取食植物中共占 63 %~76 %,占可识别植物碎片的 7011 %~8716 % (图3),一年内普氏原羚取食植物物种数有较大差异(图 4) 。8 月,普氏原羚采食 48 种植物,占当地植物种类的 6018 %,占普氏原羚采食所有植物种类的96 %,但此期普氏原羚不采食狼毒和马蔺。从11 月开始普氏原羚食物中出现狼毒和马蔺,狼毒和马蔺在食物中所占比例逐月上升,到 3 月达到最大值: 狼毒占 717 %,马蔺占 416 %。6 月,这两种植物在普氏原羚食物中消失。5 月,食物中沙蒿所占比例为 1318 %。碱蒿在 3 月曾占到食物组成的1816 %,5 月为1115 %,其它月份保持较低的比例;芨芨草是普氏原羚的基本食物,草青期氏原羚食物中芨芨草比例保持在 10 %左右。草黄期,普氏原羚对芨芨草的采食量约为草青期的倍。对普氏原羚的食物组成数据为正态分布 (K检验 , P 均大于 0105),草青期与草枯黄期普氏羚的食物组成差异极显著 (配对 t 检验 , P<0101)。
2.4 采食地
从 1998 年 8 月到 1999 年 8 月,我们共遇见普氏原羚 63 群,376 只次。普氏原羚选择芨芨草草原以及食物丰富度高、隐蔽条件好和人类干扰少的地方作为采食生境 (表 3) 。草地围栏影响普氏原羚的采食生境选择,普氏原羚在距草地围栏 1 km内的生境采食,几乎不在围栏内采食, 距围栏大于1 km则接近随机选择。普氏原羚不在沙丘中采食,而在近沙丘处采食。